Participación de los mediadores de la respuesta inmune inflamatoria en la resorción del hueso Alveolar durante la Periodontitis crónica - Revisión de literatura
Recibido Para Arbitraje: 17/03/2013
Aceptado para Publicación: 24/06/2013
Pérez-D., M., Odontólogo, Universidad de Carabobo. Magíster en Ciencias Biomédicas mención bioquímica y biología molecular. Profesor Titular del Departamento de Ciencias Morfofuncionales de la Facultad de Odontología de la Universidad de Carabobo. De Lima, A.R., Licenciada en Bioanálisis, Universidad de Carabobo. Doctora en Ciencias Biológicas Universidad Simón Bolivar. Profesor Titular del Departamento de Morfofisiopatología de la Facultad de Ciencias de la Salud de la Universidad de Carabobo. Miembro de la Sociedad Parasitológica Venezolana.
PARTICIPACIÓN DE LOS MEDIADORES DE LA RESPUESTA INMUNE INFLAMATORIA EN LA RESORCIÓN DEL HUESO ALVEOLAR DURANTE LA PERIODONTITIS CRÓNICA - REVISION DE LITERATURA
RESUMEN:
La periodontitis es un proceso patológico caracterizado por la inflamación de los tejidos de soporte de los dientes conocido como periodonto, como consecuencia de un proceso infeccioso, que puede llevar a una destrucción crónica de estos tejidos y a la consecuente pérdida dentaria. La respuesta inmunológica desencadenada en el tejido periodontal es muy compleja y depende de la integridad de la inmunidad innata y adaptativa de los tejidos, así como de los factores de riesgo modificables y genéticos, para la resolución de la infección o la evolución de la periodontitis. En el proceso inflamatorio se sintetizan derivados eicosanoides, moléculas conocidas como especies reactivas de oxígeno (ROS) y citocinas con importante participación en la estimulación de la resorción ósea mediada por el factor de inducción de osteoclastos (RANKL) que es determinante en la activación de los osteoclastos, la osteoclastogénesis del hueso alveolar y la progresión de la enfermedad periodontal.
PARTICIPATION OF THE IMMUNE RESPONSE MEDIATORS IN INFLAMMATORY ALVEOLAR BONE RESORPTION DURING CHRONIC PERIODONTITIS - A LITERATURE REVIEW
ABSTRACT
Periodontitis is a disease process characterized by inflammation of the periodontal tissues as a consequence of an infection, which can lead to chronic destruction of these tissues and the subsequent loss of teeth. The immune response triggered by the periodontal tissue is very complex and depends on the integrity of the innate and adaptive immunity of tissues, as well as modifiable risk factors and genetic for resolution of the infection or progression of periodontitis. In the inflammatory are synthesized eicosanoids derivatives, molecules known as reactive oxygen species (ROS) and cytokines, with important role in the stimulation of bone resorption mediated by osteoclasts inducing factor (RANKL), which is crucial in the activation of the osteoclasts, alveolar bone osteoclastogenesis and periodontal disease progression.
La enfermedad periodontal crónica es una patología inflamatoria de origen infeccioso que se caracteriza por la destrucción de las estructuras de soporte del diente 1; su evolución está influenciada por factores de riesgo que determinan la respuesta y persistencia de la respuesta inmune inflamatoria contra los microorganismos patógenos 2,3. Esta enfermedad afecta entre 10 y 15% de la población mundial y es la causa principal de pérdidas dentarias4. Se ha registrado una variación en la prevalencia asociada a la raza del individuo, siendo mayor en regiones al norte de África, sur de Asia o en personas de origen mediterráneo con respecto a otras zonas de Europa. En el continente Americano, en Estados Unidos afecta entre 30-50% de la población, siendo más común en las poblaciones económicamente desfavorecidas y su prevalencia disminuye con una mayor calidad de vida 5 . En Latinoamérica la periodontitis se observa con mayor frecuencia en sus formas leves y no se distinguen diferencias significativas entre los géneros 6; en Venezuela la prevalencia de la enfermedad periodontal se presenta en un 19% de la población y presenta mayor progresión de la misma a partir de los 55 años7. La importante prevalencia de la periodontitis revela un problema de salud pública cuya frecuente evolución constituye la causa principal de pacientes adultos edéntulos.
En la periodontitis la inflamación crónica se caracteriza por la producción de prostanoides, como las prostaglandinas, citocinas y de especies reactivas de oxígeno (ROS) 8,9 que promueve la actividad destructiva del tejido, al inhibir la síntesis de colágeno, activar las metaloproteasas de la matrix (MMPs) y bloquear la actividad de los inhibidores de estas proteasas (TIMPs), así como también se estimula la resorción ósea del hueso alveolar 10. Otras moléculas como las lipoxinas y resolvinas producidas en la periodontitis controlan la magnitud y duración de la inflamación y de la pérdida ósea 11, la lipoxina A4, resolvina E1 (RvE1) y la protectina D1 estimulan la remoción de las citocinas 12; específicamente la RvE1 interactúa con el receptor del leucotrieno B4 (BLT1) en neutrófilos y con el receptor ChemR23 en monocitos para regular la función de los leucocitos durante la inflamación 13; asimismo regulan la quimiotaxis de los neutrófilos, estimulan la fagocitosis y la producción de citocinas anti-inflamatorias mediada por la proteína G acoplada a receptores. Además la resolvina E2 regula la expresión de las integrinas y del factor activador de plaquetas 14.
En la presente revisión se pretende describir los mediadores de la inflamación periodontal que participan en la resorción ósea del hueso alveolar mediada por el ligando del receptor activador para el factor nuclear ?appa B (RANK) denominado RANKL o factor de inducción de osteoclastos.
MEDIADORES DE LA INFLAMACIÓN QUE PARTICIPAN EN LA RESORCIÓN ÓSEA
Los eicosanoides son moléculas derivadas del metabolismo del ácido araquidónico, localizado en las membranas biológicas de las células del huésped. Tienen una determinante actividad en el inicio y patogénesis de la lesión inflamatoria de la periodontitis, son producidos por acción de la fosfolipasa A2 y la vía de las isoenzimas cicloxigenasas o de la vía de las lipoxigenasas. Las prostaglandinas, los tromboxanos y los leucotrienos sintetizados por las células del tejido periodontal se unen a receptores transmembrana acoplados a la proteína G de las células diana para desencadenar la expresión de genes efectores de las funciones celulares.
Se han identificado ocho subfamilias de receptores de prostaglandina: D (DP1), F (FP), I (IP), el receptor de tromboxano TX (TP) y los receptores de la prostaglandina E: EP1 (prostanoide E receptor 1), EP2, EP3 y EP4 9,15 , además se conocen isoformas TP (TPα y TPα), FP ( FPA, FPB) y ocho isoformas EP3 16,17. El reconocimiento del receptor por los eicosanoides activa vías de señalización que actúan directamente sobre receptores nucleares o interacciona con inhibidores de las proteínas intracelulares generando modificación en la actividad celular 9.
La prostaglandina E2 (PGE2) es expresada por las células del sistema inmunológico frente a lipopolisacáridos (LPS) de las bacterias Gram negativas periodonto patógenos 18; tiene actividad sobre el metabolismo óseo del hueso alveolar, lo cual se ha evidenciado con la administración de anti-inflamatorios no esteroideos que inhiben la síntesis de la PGE2 y reducen la progresión de la resorción del hueso alveolar 19. Asimismo está involucrada en todo el proceso de la inflamación, dando lugar a sus signos clásicos: edema, enrojecimiento y dolor si hay alguna complicación secundaria. Durante su síntesis los estudios han demostrado una dominante actividad de la isoenzima de la prostaglandina E sintetasa microsomal (mPGES-1) con la cooperación con el factor de crecimiento de endotelio vascular (VEGF) en la promoción de la inflamación 20.
La interacción de este prostanoide al receptor EP1 o EP2 o a los receptores con menor afinidad EP1 y EP2 21, condiciona respuestas celulares que pueden ser de naturaleza pro-inflamatoria o anti-inflamatoria. La señalización intracelular para los receptores de PGE2 difieren de acuerdo al subtipo: EP1 se acopla a la movilización de calcio intracelular, EP3 inhibe la adenilato ciclasa y a la proteína quinasa A (PKA) y los receptores EP2 y EP4 estimulan la adenilato ciclasa y a PKA 22.
En la periodontitis la PGE2 se expresa principalmente en células musculares del endotelio y en fibroblastos por inducción del factor de necrosis tumoral alfa (TNF-?) e interleucina 1 beta (IL-1α) 23,24. Se ha demostrado en los fibroblastos gingivales que la expresión de PGE2 inducida por TNF-α involucra las vías señalización de la quinasa c-Jun N-terminal (JNK) activada por mitógenos y NF kappa B (NFkB) 25, además la PGE2 puede regular la función de las células dendríticas, los linfocitos T y B y de los macrófagos, como migración de las células dendríticas, la diferenciación de células T helper (Th1 o Th2) y de las células Th17, esta última mediadas por IL-23, así también contribuye a regular la expresión de las citocinas como el TNFα en células T en diferenciación. No obstante puede suprimir la diferenciación de las células Th1 y la función de células B y estimular la acción antinflamatoria en células de la inmunidad innata como neutrófilos, monocitos y células Natural Killer (NK)26.
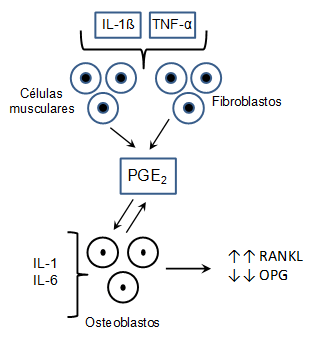
Su actividad evidencia una importante participación en la respuesta inmunológica del tejido periodontal, más aun en el hueso alveolar donde las citocinas IL-1 e IL-6 inducen en los osteoblastos la producción de PGE2, la cual actúa sobre la misma célula a través de los receptores EP4 que incrementa el AMPc intracelular y activan la expresión de RANKL y disminuye la síntesis de la osteoprotegerina (OPG) en los osteoblastos (Figura 1). Esto resulta determinante para la diferenciación de los osteoclastos y en consecuencia la resorción ósea 27,18, lo que demuestra su potencial en la osteoclastogénesis y como mediador en la patogénesis de la enfermedad periodontal.
Figura 1
Participación de las citocinas inflamatorias en la síntesis de RANKL vía derivados del ácido araquidónico.
Las citocinas también participan en la resorción ósea del hueso alveolar, su expresión es parte de la repuesta inmunológica innata y adquirida, inducida por las bacterias Gram negativas10, tales como Aggregatibacter (Actinobacillus) actinomycetemcomitans, Porphyromonas gingivalis y Tannerella forsythia que son determinantes en el desarrollo de la enfermedad periodontal1, debido a que representan patrones moleculares asociados a patógenos (PMAP) reconocidos por receptores Toll-like (TLR) y Nod-like (NLR) presentes en estas células del tejido periodontal que desencadenan la expresión de las citocinas por activación del factor de transcripción NFkB 28,29,30.
El proceso inflamatorio inducido mediante la síntesis de las citocinas que amplifican la respuesta inmunológica e inducen la producción de un conjunto de mediadores de diferenciación y activadores celulares, tales como RANKL y OPG que son determinantes en el metabolismo del hueso alveolar de lesiones periodontales activas.
En la periodontitis la IL-1, TNF-? y la interleucina 6 (IL-6) se han asociado con la osteoclastogénesis y la resorción ósea. La IL-1 es principalmente expresada por los macrófagos, fibroblastos y linfocitos T, regula la síntesis de la PGE2 y de RANKL, también estimula la proliferación de células epiteliales, fibroblastos y células endoteliales y se asocia con la activación severa de las metaloproteinasas de la matriz extracelular (MMPs) 31,32. El TNF-a está presente en altos niveles en el fluido crevicular y en el tejido periodontal donde muestra asociación con la expresión de MMPs y RANKL3,33,34, también incrementa la síntesis de PGE2 en los monocitos y macrófagos y potencia la síntesis de mediadores secundarios de la inflamación, incluyendo otros productos de la ciclooxigenasa en los linfocitos y en las células endoteliales32. La IL-6 estimula la diferenciación de los osteoclastos mediada por la expresión de RANKL vía janus kinase (JAK) /Señal transductora y activadora de la transcripción (STAT) 35 e induce la maduración de las células B para la producción de anticuerpos no específicos y estimula la síntesis de IL-110,32 que de manera indirecta inducen la expresión de RANKL en las células T 36 y en los osteoblastos3. Además, en las células del ligamento periodontal se ha demostrado que IL-6 actúa como un factor osteolítico y como mediador de la diferenciación de los osteoblastos vía factor de transcripción relacionado-Runt (Runx2) 37.
El TNF-a regula la producción de IL-1? e IL-6 10,32 y puede estimular la osteoclastogénesis directamente a través del receptor intracelular p55 presente en las células del estroma y en los precursores de los osteoblastos para mediar la expresión de RANKL38 ó indirectamente a través de la estimulación del factor estimulante de colonias de macrófagos (MCSF)39, a diferencia de IL-1 que durante la osteoclastogénesis regula la expresión de RANKL vía p38 MAPK 38, por lo tanto TNF-a e IL-1 b muestran actividad sinérgica y complementaria.
También se ha observado que pequeñas concentraciones de TNF-a estimulan la proliferación de los osteoblastos, pero concentraciones importantes de TNF-a inhiben la diferenciación de los osteoblastos40, además se ha demostrado que el factor de transcripción proteína recombinante de unión supresora de calvicie (RBPJ) tiene una participación como reguladora negativa de la osteoclastogénesis suprimiendo la actividad de TNF-a sobre la diferenciación del osteoclastos y limitando la resorción ósea en la inflamación 41. Otra citocina que se relaciona con la respuesta inflamatoria y la resorción ósea es el INF-y producido principalmente por la Th1 en presencia de bacterias Gram negativas42 probablemente debido a que participa en la regulación de la expresión de TNF-a eIL-6; no obstante otros investigadores han demostrado en condiciones in vitro que el INF-? inhibe la osteogénesis debido a que induce la degradación de la proteína adaptadora TRAF6 de RANK43 . Asimismo la interleucina 17 (IL-17) producida principalmente por los linfocitos T se ha involucrado como un inductor de la producción de RANKL 44,45. Otro grupo de mediadores de la respuesta inflamatoria de la periodontitis son los radicales libres/ROS que incluyen radicales de oxígeno libre, especies de nitrógeno y cloro con potencial antimicrobiano, generados a partir del metabolismo de los macrófagos, neutrófilos y fibroblastos, altamente reactivas capaces de generar oxidación en una variedad de biomoléculas 46 de no ser neutralizados por moléculas antioxidantes. En la periodontitis su producción es consecuencia de la respuesta a la presencia de LPS dependientes de la vía MyD88 y el incremento en los niveles de ROS se han asociado con la inducción y progreso de la enfermedad periodontal 47,48. En el tejido gingival incrementos de ROS pueden degradar los componentes de la matriz extracelular y estimular la producción del factor activador de plaquetas además de promover la trombosis intravascular 49 asimismo se ha demostrado que el efecto de ROS sobre las resorción ósea no se relaciona con el incremento de los osteoclastos, sino con aumento de RANKL sobre OPG 50.
OSTEOCLASTOGÉNESIS Y RESORCIÓN ÓSEA MEDIADA POR RANKL
La pérdida del hueso alveolar implica la activación de la osteoclastogénesis a partir de la diferenciación de células madre hematopoyéticas precursoras de los osteoclastos, como son las colonias formadoras de unidades progenitoras de granulocitos-macrófagos (CFU-GM) y las colonias formadoras de unidades progenitoras de macrófagos (CFU-M) por acción del Factor Estimulador de Colonia de macrófagos (M-CSF) que induce la expresión del receptor RANK en las células de linaje de osteoclastos durante todo su ciclo celular 51.
En la periodontitis la diferenciación de los osteoclastos tiene dos formas de estimularse: una forma indirecta como resultado de la liberación de citocinas pro-inflamatorias por las células presentes en el sitio de la infección, éstas actúan sobre los osteoblastos y los linfocitos Th1 e inducen la expresión de RANKL (Figura 1); y una segunda ruta por acción directa de LPS sobre los osteoblastos para inducir la expresión de RANKL52.
El LPS interactúa con receptores Toll-like receptor 4 (TLR4) expresado en células de linaje de monocitos, tales como células dendríticas y macrófagos dendríticas, así como en células derivadas del linaje de monocitos como los preosteoclastos y osteoclastos, activando una vía de señalización intracelular con algunos transductores de señales que desencadenan una consiguiente cascada de señalización de aguas abajo, RANK como el factor 6 receptores asociado a TNF (TRAF6), o NFkB o MAPKs 53,54.
En los preosteoclastos expuestos a LPS y a M-CSF pero no a RANKL se suprime la vía JNK y NFkB, por lo cual no se expresa al factor nuclear de células T activadas (NFATc1) que participa en la expresión de genes durante la diferenciación de los osteoclastos 55,56, pero en los preosteoclatos expuestos a LPS y RANKL se estimula la activación de NFATc1 vía JNK y NFkB en la osteoclastogénesis. En los osteoclastos maduros en presencia de RANKL, el LPS promueve la supervivencia celular mediante la activación de otros factores, Akt, NFkB, y ERK a través de MyD88 55, es decir que LPS pueden tener diferentes efectos sobre la osteoclastogénesis dependiendo de la etapa de diferenciación del preosteoclasto o de activación del osteoclasto.
Las citocinas pro-inflamatorias IL-1b, IL-6 y TNF-a secretadas por los fibroblastos, macrófagos, linfocitos TCD4+ y osteoblastos inducen la expresión de RANKL. La interacción de LPS con sus receptores CD14/TLR 4 provoca la secreción directa de estas citocinas. Sin embargo en los linfocitos T la síntesis de RANKL es indirecta de la interacción con LPS ya que requieren de una célula presentadora de antígeno (CPA) como las células dendríticas, la regulación de la expresión RANKL en los linfocitos T requiere de dos señales: de la estimulación de su receptor TCR por un antígeno presentado por un MHC-II y del receptor CD28 localizado en la membrana CPA 52. En presencia de M-CSF, TNF, IL-1 e IL-6 se inducen la síntesis de RANKL por los osteoblastos y se amplifica la secreción de M-CSF 57. En los osteoblastos la expresión de RANKL es mediada por 1,25 hidroxicolecalciferol, paratohormona, por PGE2 y por citocinas liberadas por los macrófagos 58.
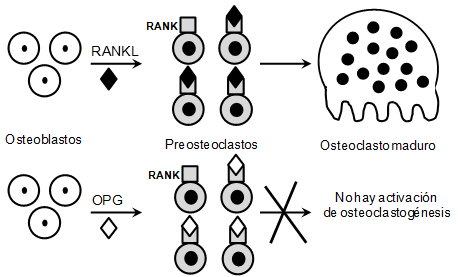
El enlace del receptor RANK expresado en los osteoclastos indiferenciados o inactivos con RANKL impulsa la osteoclastogénesis y promueve la activación y supervivencia de los osteoclastos 52. Sin embargo en los osteoblastos también se sintetiza y secreta una proteína, identificada como la osteoprotegerina (OPG), que se une específicamente a RANKL en su forma soluble o de membrana e inhibe competitivamente la interacción RANKL/RANK 59,60 (Figura 2). En condiciones en las cuales la concentración de OPG sea mayor en relación con el nivel expresado de RANKL se presenta una inhibición entre la unión entre RANKL y RANK lo que genera disminución de la velocidad e intensidad de la osteoclastogénesis y se promueva la apoptosis de osteoclastos diferenciados y activados 36,60. Por lo tanto, el equilibrio entre la expresión RANKL y OPG es esencial para la determinación de la actividad osteoclastogénica en el hueso alveolar.
Figura 2
Activación de la osteoclastogénesis mediada por RANKL.
Se ha reportado niveles superiores de OPG en tejido gingival sano, mientras que en tejido aislado de lesiones periodontales prevalecen niveles más altos de RANKL. De este modo la relación RANKL/ OPG es más elevada en pacientes con periodontitis respecto a pacientes con gingivitis o pacientes sanos, que sugiere una alta expresión de RANKL/OPG y sea la responsables de la resorción ósea periodontal 36. Existe evidencia que muestra que los fibroblastos del periodonto 61, células Th17 62 y células B 63 expresan RANKL. También los complejos inmune-IgG desarrollados por invasión bacteriana en el tejido periodontal puede promover la resorción ósea proporcionando co-estimulación de RANKL través de su unión a receptores Fc expresado en osteoclastos 36.
En el tejido periodontal los fibroblastos gingivales y los fibroblastos del ligamento periodontal también se consideran una fuente de RANKL en su forma de membrana, pero no en su forma soluble y OPG, característica similar a los osteoblastos, por lo cual se desempeña en la regulación de la homeostasis de los ligamentos periodontales que conectan hueso y el cemento 64. En los fibroblastos gingivales se ha demostrado mayor expresión de ARNm de OPG que de RANKL, inclusive se ha observado que la estimulación de IL-1b induce la expresión de OPG pero no RANKL65, lo cual se ha relacionado con la regulación de la remodelación del hueso alveolar a este nivel. Sin embargo en linfocitos T y B se ha demostrado expresión RANKL 50% y 90%, respectivamente, en el tejido con enfermedad periodontal, así también se ha descrito abundante expresión de RANKL en linfocitos T CD3+, TCD4+, T CD20+ y linfocitos B de pacientes con periodontitis crónica, pero en el tejido gingival de pacientes sanos muy pocas células sintetizan RANKL, lo que evidencia su participación en el proceso de resorción ósea 66.
Además existe evidencia que muestra una relación entre la expresión de RANKL en fibroblastos periodontales de felinos y el gen muscle segment homeobox 2 (msx2), ya que se demostró que la pérdida de expresión de este gen y disminución de los niveles de RANKL 67, probablemente se debe a la asociación de MSX2 con TNF-a vía NFk 68,69.
No obstante, la activación de los osteoclastos mediada por RANKL unido a su receptor RANK requiere de la señalización coestimuladora proporcionada por el receptor asociado a osteoclastos (OSCAR) y del receptor de activación expresado en células mieloides 2 (TREM2); OSCAR se une a un ligando específico, a un receptor Fc gamma de cadena común (FCRy) que se activa a través de la fosforilación de motivos de activación de inmunorreceptor basados en tirosina (ITAM) y regula la fosfolipasa Cy (PLCy) y la señalización de calcio en los precursores de osteoclastos lo que conduce a la inducción de la expresión de genes necesarios para la osteoclastogénesis 70.
CONCLUSIONES
Durante el proceso inflamatorio que se desarrolla en la enfermedad periodontal la osteoclastogénesis y la resorción ósea son estimuladas por la síntesis de RANKL, el cual es inducido directamente por M-CSF y TNF-? o indirectamente a través de la expresión de PGE2, ROS y otras citocinas como IL-1b, IL-6.
La activación del osteoclasto es estimulada por las bacterias periodonto patógenas que inducen un aumento en la expresión de RANKL en células del tejido gingival como los osteoblastos y fibroblastos y por células del sistema inmunológico como las células dendríticas, T y B. La interacción con el receptor RANK expresado en los preosteoclastos y osteoclastos inactivos depende de un balance entre la expresión de OPG y RANKL, en donde la OPG actúa inhibiendo la interacción de RANKL con su receptor, y por ende, la osteoclastogénesis. En presencia de bajas concentraciones de OPG, se favorece la activación del receptor RANK, quien actúa desencadenando una cascada de señalización intracelular que favorece la diferenciación de los osteoclastos, induciendo su activación y supervivencia y quienes iniciarán el proceso de resorción ósea.
REFERENCIAS BIBLIOGRÁFICAS
AAP-American Academy of Periodontology. Epidemiology of periodontal diseases. J Periodontol. 2005; 76, 1406-1419.
Laine ML, Loos BG, Crielaard W. Gene polymorphisms in chronic periodontitis. Int J Dent. 2010;1-22
Baelum V, Lopez R.Periodontal epidemiology: towards social science or molecular biology. Community Dent Oral Epidemiol. 2004 Aug;32(4):239-49.
Marchetti E, Monaco A, Procaccini L, Mummolo S, Gatto R, Tetè S, Baldini A, Tecco S, Marzo G. Periodontal disease: the influence of metabolic syndrome. Nutr Metab (Lond). 2012;9(1):88.
Mendoza C, Arteaga O, Gamonal J.Investigación Epidemiológica en Enfermedades Periodontales en América Latina .Rev Chil Period Oseoint .2006; 3(3):7-13.
Morón Borjas, Alexis. Perfil epidemiológico bucal de las etnias venezolanas. Primer reporte nacional Ciencia Odontológica [en línea] 2008, 5 (Diciembre) : [fecha de consulta: 29 de abril de 2012] Disponible en: ISSN 1317-8245
Hasturk H, Kantarci A, Goguet-Surmenian E, Blackwood A, Andry C, Serhan CN, Van Dyke TE. Resolvin E1 regulates inflammation at the cellular and tissue level and restores tissue homeostasis in vivo. J Immunol. 2007;179(10):7021-7029.
Tilley, S. L., T. M. Coffman, B. H. Koller. Mixed messages: modulation of inflammation and immune responses by prostaglandins and thromboxanes. J. Clin. Invest. 2001; 108: 15-23.
Graves DT, Oates T, Garlet GP. Review of osteoimmunology and the host response in endodontic and periodontal lesions. J Oral Microbiol. 2011;3:5304
Serhan C N. Novel Lipid Mediators and Resolution Mechanisms in Acute InflammationTo Resolve or Not? AJP.2010; 177( No. 4):1576-91.
Fredman AG, Sun YP, Kantarci A, Van Dyke TE, Luster AD, Serhan C. 2006. Apoptotic neutrophils and T cells sequester chemokines during immune response resolution through modulation of CCR5 expression. Nat. Immunol.2006;7: 1209-1216.
Akira S, Takeda K, Kaisho T. Toll-like receptors: critical proteins linking innate and acquired immunity. Nat Immunol. 2001;2:675-680.
Oh SF, Dona M, Fredman G, Krishnamoorthy S, Irimia D, Serhan CN. Resolvin E2 formation and impact in inflammation resolution. J Immunol. 2012 May 1;188(9):4527-4534.
Narumiya S, FitzGerald GA. Genetic and pharmacological analysis of prostanoid receptor function.J Clin Invest. 2001; 108:25-30.
Breyer RM, Bagdassarian CK, Myers SA, Breyer MD. Prostanoid receptors: Subtypes and signaling. Annu Rev Pharmacol Toxicol. 2001; 41:661-690.
Sugimoto Y, Negishi M, Hayashi Y, Namba T, Honda A, Watabe A, Hirata M, Narumiya S, Ichikawa A 1993 Two isoforms of the EP3 receptor with different carboxyl-terminal domains. Identical ligand binding properties and different coupling properties with Gi proteins. J Biol Chem 268:2712-2718.
Shoji M, Tanabe N, Mitsui N, Suzuki N, Takeichi O, Katono T, Morozumi A, Maeno M. Lipopolysaccharide enhances the production of nicotine-induced prostaglandin E2 by an increase in cyclooxygenase-2 expression in osteoblasts. Acta Biochim Biophys Sin (Shanghai). 2007 Mar;39(3):163-172.
Jeffcoat MK, Page R, Reddy M, Wannawisute A, Waite P, Palcanis K, Cogen R, Williams RC, Basch C.Use of digital radiography to demonstrate the potential of naproxen as an adjunct in the treatment of rapidly progressive periodontitis. J Periodontal Res. 1991 Sep;26(5):415-421.
Kamei D, Yamakawa K, Takegoshi Y, Mikami-Nakanishi M, Nakatani Y, Oh-Ishi S, Yasui H, Azuma Y, Hirasawa N, Ohuchi K, Kawaguchi H, Ishikawa Y, Ishii T, Uematsu S, Akira S,Murakami M, Kudo I. Reduced pain hypersensitivity and inflammation in mice lacking microsomal prostaglandin E synthase-1. J Biol Chem. 2004; 279:33684-95.
Sugimoto Y, Narumiya S. Prostaglandin E receptors. J Biol Chem. 2007; 282:11613-7.
Suzawa, T., C. Miyaura, M. Inada, T. Maruyama, Y. Sugimoto, F. Ushikubi, A. Ichikawa, S. Narumiya, T. Suda. 2000. The role of prostaglandin E receptor subtypes (EP1, EP2, EP3, and EP4) in bone resorption: An analysis using specific agonists for the respective EPs. Endocrinology 141: 1554-1559
Bage T, Lindberg J, Lundeberg J, Modéer T, Yucel-Lindberg T.Signal pathways JNK and NF-kappaB, identified by global gene expression profiling, are involved in regulation of TNFalpha-induced mPGES-1 and COX-2 expression in gingival fibroblasts. BMC Genomics. 2010 Apr 15;11:241.
Yucel-Lindberg T, Hallström T, Kats A, Mustafa M, Modéer T.Induction of microsomal prostaglandin E synthase-1 in human gingival fibroblasts. Inflammation. 2004;28(2):89-95.
Bage T, Kats A, Lopez BS, Morgan G, Nilsson G, Burt I, Korotkova M, Corbett L, Knox AJ, Pino L, Jakobsson PJ, Modéer T, Yucel-Lindberg T.Expression of prostaglandin E synthases in periodontitis immunolocalization and cellular regulation. Am J Pathol. 2011;178(4):1676-88.
Ricciotti E, FitzGerald GA. Prostaglandins and Inflammation.Arterioscler Thromb Vasc Biol. 2011 May;31(5):986-1000.
Inada M, Matsumoto C, Uematsu S, Akira S, Miyaura C.Membrane-bound prostaglandin E synthase-1-mediated prostaglandin E2 production by osteoblast plays a critical role in lipopolysaccharide-induced bone loss associated with inflammation. J Immunol. 2006 Aug 1;177(3):1879-85.
Murthy MB.Osteoimmunology - Unleashing the concepts. J Indian Soc Periodontol. 2011;15(3):190-198.
Kitaura H, Sands MS, Aya K, Zhou P, Hirayama T, Uthgenannt B, Wei S, Takeshita S, Novack DV, Silva MJ, Abu-Amer Y, Ross FP, Teitelbaum SL. Marrow stromal cells and osteoclast precursors differentially contribute to TNF-alpha-induced osteoclastogenesis in vivo. J Immunol. 2004 Oct 15;173(8):4838-4846.
Abu-Amer, Y., F. P. Ross, J. Edwards, S. L. Teitelbaum. Lipopolysaccharide-stimulated osteoclastogenesis is mediated by tumor necrosis factor via its p55 receptor. J. Clin. Invest. 1997;100(6):1557-65
Guzeldemir E, Gunhan M, Ozcelik O, Tastan H.Interleukin-1 and tumor necrosis factor-alpha gene polymorphisms in Turkish patients with localized aggressive periodontitis. J Oral Sci. 2008;50(2):151-159.
Okada H, Murakami S.Cytokine expression in periodontal health and disease. Crit Rev Oral Biol Med. 1998;9(3):248-266.
Graves D.T., Cochran D. The contribution of interleukin-1 and tumor necrosis factor to periodontal tissue destruction. J Periodontol. 2003;74:391-401.
Garlet GP, Martins W Jr, Fonseca BA, Ferreira BR, Silva JS.Matrix metalloproteinases, their physiological inhibitors and osteoclast factors are differentially regulated by the cytokine profile in human periodontal disease. J Clin Periodontol. 2004 Aug;31(8):671-679.
Hashizume M, Hayakawa N, Mihara M.IL-6 trans-signalling directly induces RANKL on fibroblast-like synovial cells and is involved in RANKL induction by TNF-alpha and IL-17. Rheumatology (Oxford). 2008 Nov;47(11):1635-1640.
Kajiya M, Giro G, Taubman MA, Han X, Mayer MP, Kawai T.Role of periodontal pathogenic bacteria in RANKL-mediated bone destruction in periodontal disease. J Oral Microbiol. 2010;2:5532.
Iwasaki K, Komaki M, Mimori K, Leon E, Izumi Y, Ishikawa I. IL-6 induces osteoblastic differentiation of periodontal ligament cells. J Dent Res. 2008 Oct;87(10):937-942.
Gilbert L, He X, Farmer P, Boden S, Kozlowski M, Rubin J, Nanes MS.Inhibition of osteoblast differentiation by tumor necrosis factor alpha. Endocrinology. 2000 Nov;141(11):3956-3964.
Zhao B, Grimes SN, Li S, Hu X, Ivashkiv LB. TNF-induced osteoclastogenesis and inflammatory bone resorption are inhibited by transcription factor RBP-J. J Exp Med. 2012 Feb 13;209(2):319-334.
Garlet TP, Coelho U, Repeke CE, Silva JS, Cunha Fde Q, Garlet GP.Differential expression of osteoblast and osteoclast chemmoatractants in compression and tension sides during orthodontic movement..Cytokine. 2008 Jun;42(3):330-33
Takayanagi H, Ogasawara K, Hida S, Chiba T, Murata S, Sato K, Takaoka A, Yokochi T, Oda H, Tanaka K, Nakamura K, Taniguchi T.T-cell-mediated regulation of osteoclastogenesis by signalling cross-talk between RANKL and IFN-gamma.Nature. 2000 Nov 30;408(6812):600-60
Kotake S, Udagawa N, Takahashi N, Matsuzaki K, Itoh K, Ishiyama S, Saito S, Inoue K, Kamatani N, Gillespie MT, Martin TJ, Suda T. IL-17 in synovial fluids from patients with rheumatoid arthritis is a potent stimulator of osteoclastogenesis. J Clin Invest. 1999 May;103(9):1345-1352.
Sato K, Suematsu A, Okamoto K, Yamaguchi A, Morishita Y, Kadono Y,et al. Th17 functions as an osteoclastogenic helper T cell subset that links T cell activation and bone destruction. J Exp Med. 2006; 203:2673-2682.
Dahiya P, Kamal R. Gupta R, Puri A.Oxidative stress in chronic periodontitis. Chronicles of young scientists.2011;2 (4):179-181.
Tsai CC, Chen HS, Chen SL, Ho YP, Ho KY, Wu YM, Hung CC.Lipid peroxidation: a possible role in the induction and progression of chronic periodontitis.J Periodontal Res. 2005 Oct;40(5):378-384
Konopka T, Król K, Kope? W, Gerber H. Total antioxidant status and 8-hydroxy-2'-deoxyguanosine levels in gingival and peripheral blood of periodontitis patients. Arch Immunol Ther Exp (Warsz). 2007 Nov-Dec;55(6):417-22
Van Dyke TE, Serhan CN. Resolution of inflammation: a new paradigm for the pathogenesis of periodontal diseases. J Dent Res. 2003;82(2):82-90.
Kondo H, Takeuchi S, Togari A. ?-Adrenergic Signaling Stimulates Osteoclastogenesis via Reactive Oxygen Species. Am J Physiol Endocrinol Metab. 2013;304(5):E507-15.
Amcheslavsky A, Bar-Shavit Z. Toll-like receptor 9 ligand blocks osteoclast differentiation through induction of phosphatase. J Bone Miner Res. 2007;22:1301-1310.
Morgan L Philippe.Activation de l'ostéoclasie par les endotoxines bactériennes au coursdes maladies parodontales. Medecine/Sciences. 2006;22: 614-619.
Hans M, Hans VM.Toll-like receptors and their dual role in periodontitis: a review. J Oral Sci. 2011;53(3):263-271.
Arita, M., T. Ohira, Y. P. Sun, S. Elangovan, N. Chiang, C. N. Serhan. Resolvin E1 selectively interacts with leukotriene B4 receptor BLT1 and ChemR23 to regulate inflammation. J. Immunol. 2007;178: 3912-3917.
Liu D, Xu JK, Figliomeni L, Huang L, Pavlos NJ, Rogers M, et al. Expression of RANKL and OPG mRNA in periodontal disease: possible involvement in bone destruction. Int J Mol Med. 2003;11:17-21.
Takayanagi H. The role of NFAT in osteoclast formation. Ann NY Acad Sci 2007; 1116: 227-237.
Kikuchi T, Matsuguchi T, Tsuboi N, et al. Gene expression of osteoclast differentiation factor is induced by lipopolysaccharide in mouse osteoblasts via Toll-like receptors. J Immunol 2001 ;166 : 3574-3579.
Singh PP, van der Kraan AG, Xu J, Gillespie MT, Quinn JM. Membrane-bound receptor activator of NF?B ligand (RANKL) activity displayed by osteoblasts is differentially regulated by osteolytic factors. Biochem Biophys Res Commun. 2012 May 25;422(1):48-53.
Simonet WS, Lacey DL, Dunstan CR, Kelley M, Chang MS, Lüthy R, Nguyen HQ, Wooden S, Bennett L, Boone T, Shimamoto G, DeRose M, Elliott R, Colombero A, Tan HL, Trail G, Sullivan J, Davy E, Bucay N, Renshaw-Gegg L, Hughes TM, Hill D, Pattison W, Campbell P, Sander S, Van G, Tarpley J, Derby P, Lee R, Boyle WJ. Osteoprotegerin: a novel secreted protein involved in the regulation of bone density. Cell. 1997 Apr 18;89(2):309-319
Yasuda H, Shima N, Nakagawa N, Yamaguchi K, Kinosaki M, Mochizuki S, Tomoyasu A, Yano K, Goto M, Murakami A, Tsuda E, Morinaga T, Higashio K, Udagawa N, Takahashi N, Suda T.Osteoclast differentiation factor is a ligand for osteoprotegerin/osteoclastogenesis-inhibitory factor and is identical to TRANCE/RANKL Proc Natl Acad Sci U S A. 1998 Mar 31;95(7):3597-3602.
Bloemen V, Schoenmaker T, de Vries TJ, Everts V.Direct cell-cell contact between periodontal ligament fibroblasts and osteoclast precursors synergistically increases the expression of genes related to osteoclastogenesis. J Cell Physiol. 2010 Mar;222(3):565-573.
Dutzan N, Gamonal J, Silva A, Sanz M, Vernal R.Over-expression of forkhead box P3 and its association with receptor activator of nuclear factor-kappa B ligand, interleukin (IL) -17, IL-10 and transforming growth factor-beta during the progression of chronic periodontitis. J Clin Periodontol. 2009 May;36(5):396-403.
Han X, Lin X, Seliger AR, Eastcott J, Kawai T, Taubman MA.Expression of receptor activator of nuclear factor-kappaB ligand by B cells in response to oral bacteria.Oral Microbiol Immunol. 2009 Jun;24(3):190-196
Kanzaki H, Chiba M, Shimizu Y, Mitani H. Dual regulation of osteoclast differentiation by periodontal ligament cells through RANKL stimulation and OPG inhibition. J Dent Res. 2001;80:887-891.
Hormdee D, Nagasawa T, Kiji M, Yashiro R, Kobayashi H, Koshy G, et al. Protein kinase-A-dependent osteoprotegerin production on interleukin-1 stimulation in human gingival fibroblasts is distinct from periodontal ligament fibroblasts. Clin Exp Immunol. 2005;142:490-497.
Kawai T, Matsuyama T, Hosokawa Y, Makihira S, Seki M, Karimbux NY, et al. B and T lymphocytes are the primary sources of RANKL in the bone resorptive lesion of periodontal disease. Am J Pathol. 2006;169:987-998.
Booij-Vrieling HE, Ferbus D, Tryfonidou MA, Riemers FM, Penning LC, Berdal A, Everts V, Hazewinkel HA.Increased vitamin D-driven signalling and expression of the vitamin D receptor, MSX2, and RANKL in tooth resorption in cats. Eur J Oral Sci. 2010 Feb;118(1):39-46.
Lee HL, Yi T, Woo KM, Ryoo HM, Kim GS, Baek JH. Msx2 mediates the inhibitory action of TNF-alpha on osteoblast differentiation.Exp Mol Med. 2010 Jun 30;42(6):437-445.
Lai CF, Shao JS, Behrmann A, Krchma K, Cheng SL, Towler DA.TNFR1-activated reactive oxidative species signals up-regulate osteogenic Msx2 programs in aortic myofibroblasts.Endocrinology. 2012 Aug;153(8):3897-3910.
Koga T, Inui M, Inoue K, Kim S, Suematsu A, Kobayashi E, et al. Costimulatory signals mediated by the ITAM motif cooperate with RANKL for bone homeostasis. Nature. 2004;428:758-763.