Apoptosis mediada por Ceramidas en la Carcinogenesis inducida de Glándula Parótida
Recibido para arbitraje: 11/12/2009
Aceptado para publicación: 16/04/2010
Ana Maria Zarate: Dra. En Ciencias de la Salud. Profesor adjunto Cátedra de Biología Celular. Facultad de Odontología. Universidad Nacional de Córdoba. e-mail: [email protected]. Dirección: Haya de la Torre s/n. Ciudad Universitaria. Córdoba. Argentina
Mabel Noemí Brunotto: Dra. En Ciencias de la Salud. Profesor Titular Cátedra de Biología Celular. Facultad de Odontología. Universidad Nacional de Córdoba. Argentina.
Dra. Alicia Inés Malberti: Dra. En Ciencias Biológicas. Profesor asistente Cátedra de Histología y Embriología. Facultad de Odontología. Universidad Nacional de Córdoba. Argentina.
Dra. Reyna Olga Calderón: Dra. En Ciencias Químicas. Profesora consulta. Facultad de Ciencias Médicas. Universidad Nacional de Córdoba. Argentina.
RESUMEN
La apoptosis es un proceso relacionado con el cáncer ya que las células malignas evaden los mecanismos apoptóticos. La parótida es el sitio más común de tumores de glándulas salivales y solo el 15 % es detectado en estadios tempranos. Conocer que ocurre con las moléculas involucradas en la apoptosis y utilizarlas como biomarcadores, resultaría de importancia para aplicar nuevas terapias y prevención. En este trabajo estudiamos alteraciones tempranas de la vía apoptótica inducida por Fas, mediada por ceramidas en tumorogénesis experimental. Se utilizaron ratas inyectadas con 9, 10- dimethyl 1, 2 benzanthracene (DMBA), en cada glándula parótida. Las muestras fueron analizadas a 7; 30 y 150 días post tratamiento. Se realizó coloración de (H/E), inmunocitoquímica para Fas, determinación de ceramidas por cromatografía e capa delgada de alto rendimiento, "patrón en escalera" de ADN en gel de agarosa y secuenciación del los exones 3 y 4 del gen fas. A 7 días se presentaron cambios histológicos leves y una mutación en el exón 4. No se detectó la presencia de ceramidas. A 30 y 150 días hubo cambios de tipo tumorogénicos y necrosis, mutaciones en el exón 4 de Fas y presencia de ceramidas. A ningún tiempo el patrón en escalera de ADN demostró apoptosis. A los 7 días no se producen cambios suficientes par inducir la apoptosis, mientras que a 30 y 150 días se estaría formando el complejo Fas-Fas L con aumento de las ceramidas , pero las mutaciones del exón 4 podrían estar modificando la proteína Fas a nivel de su dominio de muerte impidiendo así la transducción de la señal de apoptosis.
ABSTRACT
Apoptosis is a process linked to cancer because malignant cells evade apoptotic mechanisms. The parotid gland is the most common site of salivary gland tumors and only 15% is detected in early stages. Know what happens to the molecules involved in apoptosis and use as biomarkers, it would be important to apply new therapies and prevention. In this paper we study early alterations of the Fas-induced apoptotic pathway mediated by ceramide in experimental tumorigenesis male rats were used, injected with 9, 10 - dimethyl 1, 2 benzanthracene (DMBA) in each parotid gland. The samples were analyzed at 7, 30 and 150 days after treatment. Staining H / E, was performed immunocytochemistry for Fas, determination of ceramides by High performance thin layer chromatography ( HPTLC), ladder of DNA in agarose gel and sequencing of exons 3 and 4 of the fas gene. A 7 days it showed mild histological changes and a mutation in exon 4. Not ceramides were detected . At 30 and 150 days we found tumourigenic changes and necrosis type, mutations in exon 4 of Fas and the presence of ceramides. At no time DNA ladder showed apoptosis. After 7 days no change enough torque to induce apoptosis, while at 30 and 150 days would be forming the complex Fas-Fas L with increase of ceramides, but mutations of exon 4 could be changing the Fas protein level of his death domain thereby preventing the signal transduction of apoptosis.
La apoptosis es un proceso innato y evolutivamente conservado, en el cual las células se inactivan, se desensamblan y degradan su propia estructura y componentes de manera característica (1). Durante la apoptosis se produce la fragmentación del ADN de la célula apoptótica mientras se mantiene la integridad de las membranas. En este proceso podemos distinguir varias fases: D1 donde se produce los mecanismos moleculares que inician el fenómeno; fase F en la que se fragmenta el ADN; fase D2 en la cual se produce la destrucción nuclear y del citoplasma y los restos celulares son fagocitados por macrófagos (2). La apoptosis es llevada a cabo por una gran familia de enzimas proteasas, llamadas caspasas. (3). Los estímulos que generan la apoptosis son diversos, y generan señales intra o extracelulares que serán detectadas por las células. Las señales intracelulares producen la liberación del citocromo C del espacio intermembranoso de la mitocondria (4); las señales extracelulares provocan la unión de los ligandos a sus respectivos receptores de superficie celular, conocidos como receptores de muerte (DR). Los DR pertenecen a la familia de proteínas del Factor de Necrosis Tumoral (TNF), caracterizadas por presentar dominios extracelulares ricos en cisteínas. Dentro de esta gran familia el receptor mejor caracterizado es Fas (CD95 ó Apo-1) ya que es considerado un regulador clave en el proceso de apoptosis (5). La mayoría de las células humanas expresan la proteína Fas. El receptor Fas (APO/-1/CD-95) es una glucoproteína transmembrana rica en cisteína de 48 kilodaltons (kDa). Como todos los DR lleva una banda de 80 aminoácidos altamente conservada, que se conoce con el nombre de dominio de muerte (DD), ubicada en su porción citoplasmática, el cual es fundamental para el inicio de la apoptosis. (6). Para que la apoptosis ocurra se requiere de un dominio de muerte intacto en la porción citoplasmática de Fas. Mutaciones en el DD abortan la inducción de la apoptosis y se encuentran naturalmente en ratones y en pacientes con síndrome linfoproliferativo autoinmune (7,8). El ligando Fas-L (CD95-L ó Apo/1-L) también pertenece a la familia de TNF y es expresado tanto en la membrana celular (mFas -L), en cuyo caso se trata de una proteína transmembrana de tipo II, como en forma soluble (s Fas-L). La forma soluble, Fas-L, se deriva de mFas-L por el clivaje de las metaloproteinasas. (9).
Aunque el sistema Fas-FasL fue originalmente descrito en el contexto de la apoptosis en linfocitos, ha sido detectado también en otros tejidos tales como cerebro, testículo, ojo, mama y en neoplasias malignas (10). Su expresión también ha sido determinada en órganos humanos con enfermedades inflamatorias crónicas, tales como artritis reumatoide y colitis ulcerativas y en glándulas salivales de pacientes con Síndrome de Sjögren (11).
El sistema Fas-FasL ha sido relacionado con las ceramidas. (12). Las ceramidas, son lípidos formados por esfingosina y un ácido graso en unión amídica (13); se generan como consecuencia de numerosos estímulos tanto apoptóticos como de estrés (14) y han sido propuestas como mensajeros para eventos tan diversos como la diferenciación, senescencia, proliferación, arresto del ciclo celular y apoptosis (15). Se considera que la principal vía para la producción de ceramida es la hidrólisis de la esfingomielina, por la acción de la enzima esfingomielinasa (16).
Kolesnik (12) propuso un modelo, en el cual la esfingomielinasa ácida genera ceramida, formando balsas (rafts ) de membrana, los cuales serían críticos para la oligomerización de receptores y el ensamblado de estructuras complejas de señalización . La interacción de la esfingomielinasa ácida con los receptores dentro de estas balsas ha sido reportada para moléculas tales como p75, CD40 y Fas. (17,18, 19).
El modelo de activación de la vía apoptótica por estímulos externos, captados por receptores de membrana y mediado por ceramidas propuesto por Kolesnik (12) indica que la unión de los Fas (CD95/Apo-1) triméricos preformados, de una célula blanco, a FasL activa pequeñas cantidades de caspasa 8 dentro del citoplasma de la célula, que no obstante son suficientes para inducir la translocación de la esfingomielinasa ácida, desde su localización intracelular, a la capa externa de la membrana plasmática, que es rica en esfingomielina, produciendo allí el clivage de la misma en ceramida y fosforilcolina. Se induce de este modo la agrupación de los Fas en balsas (rafts) de membrana ricas en ceramidas que probablemente son el resultado de la segregación lateral de las mismas, después de la unión de los receptores de superficie a sus ligandos o a anticuerpos agonistas. La ceramida, que se autoasocia, inicia la coalescencia de balsas (rafts) ricos en ceramida formando grandes parches y plataformas. Fas unido a Fas L pueden entrar y concentrarse dentro de estas plataformas, y lo hacen para reclutar a la proteína adaptadora FADD, (por una interacción homotípica con su DD), formando el complejo inductor de muerte (DISC). La proteína FADD recluta entonces la procaspasa 8 al DISC (a través de una nueva interacción homotípica, entre los dominios efectores de muerte). La procaspasa 8 se cliva en caspasa 8 y activa la caspasas efectoras 3,6 y 7 y comienza así la ejecución del proceso de apoptosis (17, 20). Se han realizado estudios en los cuales se demuestra que las modificaciones en esta vía pueden contribuir a la patogénesis del cáncer (21). El objetivo de este trabajo fue estudiar, utilizando un modelo experimental de inducción de la tumorogénesis en parótida, modificaciones tempranas en la vía de apoptosis inducida por el sistema Fas-Fas-L relacionado a ceramidas presentes en la membrana plasmática.
MATERIAL Y MÉTODOS
El protocolo utilizado en esta investigación, sigue los lineamientos de National Institutes of Health (NIH) para el uso de animales de laboratorio, y fue revisado y aprobado por el Comité de Ética en Investigación del Hospital Pediátrico del Niño Jesús de la Provincia de Córdoba, Argentina. Se utilizaron 80 ratas machos, de la variedad Wistar, de dos meses de edad con peso aproximado de 180 a 200g, alimentadas ad libitum con dieta balanceada y agua, mantenidas con temperatura controlada de 24Cº y con ciclos de luz / oscuridad de 12:12 h. Los animales se separaron en dos grupos: a) experimental (n=40), al que se le indujo la carcinogénesis, con inyección de 50?l de una solución al 0.5 % de 9, 10- dimethyl 1, 2 benzanthracene (DMBA) (SIGMA ALDRICH Chemical Industries, USA) disuelto en acetona, en cada glándula parótida, expuesta por cirugía, y b) control (n=40), al que se le inyectó la misma cantidad de solución fisiológica en ambas glándulas parótidas, expuestas por cirugía.
Inducción de la tumorogénesis: los animales, de ambos grupos, fueron anestesiados por vía intraperitoneal con una inyección de solución de 1.28mg Xilazine / 8mg Ketamine / 100g de peso corporal. (Bayer, Argentina y Ketalar, Parker Davis, respectivamente). Posteriormente se les realizó una incisión en la superficie ventral del cuello, para la exposición de las glándulas parótidas (14). Se tomaron muestras de las glándulas a los 7, 30 y 150 días después de la inyección, se pesaron en balanza de precisión y siempre la glándula parótida derecha fue procesada y utilizada para microscopía óptica y la izquierda para estudios bioquímicos y moleculares.
Estudios histológicos: Las muestras de glándulas parótidas se fijaron en formol al 10%, tamponado con acetil piridina clorhídrico, pH 7, se incluyeron en parafina, se colorearon con Hematoxilina / Eosina (H/E) se observaron con microscopio óptico Olimpus BX50 y las imágenes fueron capturadas con cámara de video SONY y software Image-ProPlus, 1999.aa.
Determinación de ceramida: Se realizó por medición de la actividad de la enzima esfingomielinasa ácida utilizando un sustrato fluorescente (NBD-sphingolielin) y cromatografía en capa fina de alta resolución (HPTLC) (22).
Para ello se diluyó el sustrato fluorescente (6(N-(7 nitrobenz-2-oxa-1,3 diazol-4-yl) amino)-(hexanoyl)sphingosyl-phosphocholine) NBD C6 sphingomyelin) (Molecular Probes Invitrogen) ( NBD-Sh), en 1ml de dimetilsulfóxido. De esta solución se tomó una alícuota de 1,5 ?l a la que se le agregó 2 ml de buffer de reacción ácido (0,2 gr. de acetato de sodio; 0,003 gr. de EDTA y 10 ml de agua) para preparar una solución madre de NBD-Sh. Se realizó un homogenato de tejido glandular, a los tiempos de tratamiento estudiados, utilizando 0,030 gr. de glándula y 500 ?l de buffer de reacción. Se tomaron alícuotas apropiadas de estas suspensiones para la determinación de proteínas por técnica de Lowry (23).Cada homogenato (conteniendo 300 ?g de proteínas), se llevó a volumen final de 200 ?l con buffer ácido de reacción, se le agregó 100 ?l de la solución madre de NBD-esfingomielina y se incubó a 37º C en baño termostatizado, durante una hora. La reacción fue detenida por el agregado de 1000 ?l de cloroformo: metanol, 2:1 (V/V). Las muestras se centrifugaron a 2000 rpm durante 10´, se desechó la fase superior y la fase inferior fue llevada a sequedad bajo una corriente de N2. El sedimento se retomó en 25 ?l de cloroformo y tanto las muestras (25 ?l) como los estándares (5 ?l) fueron sembradas en placa finas de sílica gel utilizando como solvente para la corrida cromatografíca el sistema de solvente constituido por cloroformo: metanol: agua 65:25:4(v/v). Los estándares utilizados fueron: monoglicérido, diglicérido, triglicérido, cerebrósido, NBD-sphingomielin, sulfátido y ceramida. La fluorescencia emitida por los compuestos separados cromatográficamente fue visualizada después de la excitación con luz UV o bien mediante el revelado de fosfolípidos con S04Cu-SO4H2. Las imágenes fueron tomadas con una cámara digital Cannon Eos Rebel T1i1. La fluorescencia emitida fue cuantificada por densitometría después de la digitalización de la imagen utilizando el programa Image Pro-Plus, 1999.
Inmunocitoquímica: Se realizó mediante la técnica de inmunoperoxidasa streptavidina biotina. Las muestras fueron desparafinadas con xilol, deshidratadas en alcohol y lavadas con agua destilada. Luego fueron incubadas durante 2 h. a 4ºC en cámara húmeda con el anticuerpo policlonal Fas (Santa Cruz Biotechnology, USA), en concentración 1/200 y posteriormente durante 2 h con el anticuerpo secundario, consistente en una mezcla de suero anti ratón, anti conejo y anticabra biotinizado (1:200). Finalmente se le adicionó el complejo streptavidina/peroxidasa y la marcación se reveló con tetrahidroclorato de diaminobenzidina (DAKO, USA). Las muestras coloreadas se deshidrataron y se montaron con Entellan (Merck, Darmstadt, Alemania). Estudios controles se realizaron siguiendo el mismo protocolo, pero omitiendo el anticuerpo primario (24). Los cortes se observaron fueron analizadas por microscopía de epifluorescencia con un microscopio Olimpus BX50. Las imágenes fueron capturadas con cámara de video SONY y software Image-ProPlus, 1999.aa
Determinación de "patrón en escalera". Se utilizaron 30 mgr. de glándula, con características tumorales a los tiempos 7, 30 y 150 días según el protocolo de Gong J. (25). La muestra se colocó en PBS 4x, se cortó en secciones de aproximadamente 1mm3, se lavó con PBS y se colocó en medio de cultivo DMEM (Minimal Essential Médium) adicionado con pronasa 0,01% (10 ?mg/100ml) (Sigma Chemical Co) y se incubó a 37º C durante 45 minutos con agitación mágnética. Posteriormente la suspensión fue colada y el filtrado se centrifugó a 2000 rpm durante 10 minutos. Se descartó el sobrenadante y se repitió el lavado con igual volumen de PBS. El pellet del último lavado se resuspendió en 1ml. de PBS y la concentración celular del mismo se determinó después de contar las células en una alícuota apropiada mediante el microscopio óptico. La extracción diferencial de ADN de bajo peso molecular se realizó fijando la muestra en etanol 70%, 1:5 (muestra-etanol) y se guardó a -20º C.en buffer citrato. Posteriormente la muestra se centrifugó a 12000 rpm durante 5´, se eliminó el etanol (sobrenadante) y se dejó secar durante 30 minutos a temperatura ambiente. Se volvió a centrifugar a 12000 rpm durante 5 minutos y se recogió el sobrenadante en tubos nuevos.(26) A cada tubo se le colocó proteinasa K (5?l) a 50ºC, durante 30 minutos. Se volvió a centrifugar a 10000 rpm durante 5 minutos y se recuperó el sobrenadante. Se colocó 1vol de isopropanol, precipitando así el ADN. Se centrifugó a 10000 rpm y se descartó el sobrenadante. El precipitado de ADN se lavó con etanol 70 %, se centrifugó y se dejó secar. Se resuspendió en buffer TE 1x (100 ?l). La concentración de ADN en ?g/ml se midió espectrofotométricamente a 260 nm de longitud de onda con UV. Posteriormente se corrió en gel de agarosa 2%. A 50V. , colocándose alícuotas necesarias para igualar la concentración de ADN en cada calle (.2 x 106 células /500 ?l). Como controles, se utilizaron glándulas normales y tumores con apoptosis confirmada por citometría de flujo y microscopía óptica.
Aislamiento de ADN. El ADN genómico fue aislado de muestras de 10 mg de parótida, con características macroscópicas tumorogénicas, utilizando un kit de purificación (Wizard Genomic DNA purification kit-Promega). La cantidad de ADN obtenido fue determinado por la relación de la absorvancia . El índice de esta relación > 1.70 indicó un aceptable grado de pureza. Estas preparaciones fueron utilizadas para la amplificación por Reacción en Cadena de la Polimerasa (PCR).
Las mutaciones fueron detectadas utilizando PCR. Se amplificaron las regiones 8903 a 9040 (exón 3) y 10064 a 10172 (exón 4) del gen fas , número de acceso NW_047565 de Genbank y las regiónes 7 a 412 (exón 1), región 1099 a 1144 (exón 2), región 5091 a 5147 (exón 3) y región 6154 a 6548 (exón 4) del gen fas-L de Rattus norvegicus, publicadao en Genbank, con número de acceso NW_005112) (Figura 4.2.). Los primers utilizados figuran en tablas 1 y 2. La reacción de PCR fue realizada a un volumen final de 50 ?l. La amplificación se llevó a cabo en un termociclador (BIO-Rad iCycler) de acuerdo al siguiente protocolo: 10 min a 95Cº, 40 ciclos de : 1 min a 95Cº , 1 min a 62 Cº y 2 min a 72Cº p; y una extensión final a 72Cº por 10 min. Se utilizaron aproximadamente 10 ng de DNA problema. Los productos de PCR fueron fraccionados en geles de agarosa 0.8% en Tris/Borato/EDTA (TBE) y teñidos con Bromuro de Etidio. Se utilizó un marcador de ADN Ladder de 0.5 a 12.2 bp (Cien Marker- Promega USA). Los productos amplificados por PCR del gen Fas fueron purificados en columna y secuenciados en Macrogen INC (Seúl, Corea).
Tabla 1
Cebadores utilizados para la amplificación de los exones 3 y 4 del gen Fas de rata. Tabla 2
Cebadores utilizados para la amplificación del gen Fas L de rata. Análisis estadísticos: La comparación del valor medio de las concentraciones de las densidades ópticas de esfingomielina y ceramidas, a los diferentes tiempos estudiados, fue evaluado mediante prueba de ANAVA bifactorial a efectos fijos con interacción y contrastes ortogonales para comparaciones post hoc, fijando p<0.05 para diferencias estadísticamente significativas. (27). En tanto, la inmunomarcación de Fas en los tiempos estudiados fue analizada con un prueba no paramétrica de Wilcoxon, considerando inmunomarcación negativa a un 5% de área marcada por área estándar considerada, fijando un valor p<0.05 para diferencias significativas entre los porcentajes de los grupos control y problema en cada tiempo estudiado. Para el análisis de los datos se utilizó el software Infostat, versión profesional 2007.
RESULTADOS
Todos los animales inyectados con DMB, presentaron una induración palpable a los 30 y 150 días post-inyección, en el lugar ocupado por la parótida. Después de realizar la incisión quirúrgica, no se observaron cambios macroscópicos en las glándulas de animales de 7 días de tratamiento; dichos cambios solo fueron observados en forma consistente en animales con 30 y 150 días post-tratamiento. Estos cambios estuvieron representados por una masa dura e hipertrofiada y desorganización local de la zona glandular que dificultó la ubicación de la glándula. En la región adyacente a la dicha alteración macroscópica se observó aumento de la vascularización, fragilidad capilar y ganglios hipertrofiados. No se observaron modificaciones en los animales del grupo control.
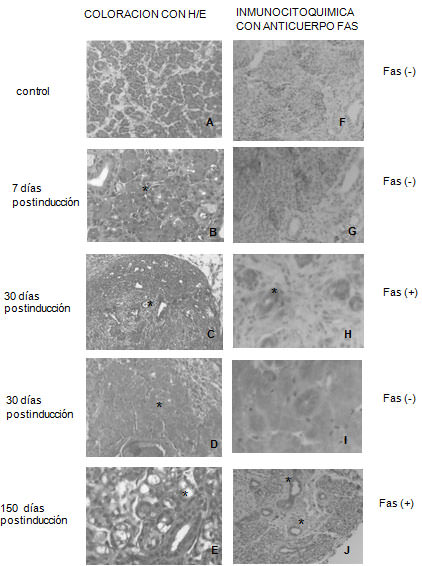
Histología de las glándulas (Figura 1 A a E): En las parótidas del grupo experimental del tiempo 7 días postinducción con DMBA, tratadas con la coloración de (H/E), presentaron cambios histológicos leves: sectores aislados con vacuolas en el citoplasma de las células acinares y ductales. Se observaron conductos intralobulillares con duplicación de capas celulares.
A los 30 días postinducción, se presentó crecimiento tumoral de tipo quístico, con islotes de células epiteliales atípicas que se encontraron infiltrando el tejido conjuntivo próximo a la glándula. También fue posible observar acúmulos de numerosos conductos intralobulillares de diámetro pequeño sin aparente presencia de acinos con núcleos que presentaron diferentes grados de coloración. Se observaron frecuentes zonas con gran eosinofilia citoplasmática, sin tinción nuclear y con conservación de los límites celulares, lo que hace suponer la existencia de necrosis por coagulación.
A los 150 días, con H/E se observaron vacuolas en células acinares, con conductos no degenerados pero con hiperplasia epitelial ductal. Las glándulas parótidas de los animales controles examinados no presentaron modificaciones histológicas
Inmucitoquímica para la proteína Fas. (Figura 1F a J) La proteína Fas demostró inmunomarcación (-), tanto en los animales controles como en los de 7 días postinducción con DMBA. A 30 días postinducción 20% de los conductos de la glándula parótida expresaron inmunomarcación (+) para Fas ; la zona de necrosis apareció con inmunomarcación (-) . A los 150 días, postinducción, el 70% de los conductos aparecieron inmunomarcados positivamente.
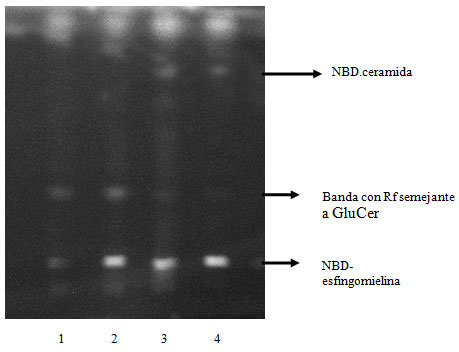
Ceramidas. (Figura 2). Bajo las condiciones de este ensayo de fluorescencia de alta precisión (HPTLC) la esfingomielina y la ceramida aparecieron como bandas fluorescentes con RF de 0.15 y 0.80 respectivamente. Tanto en los controles como en el tiempo 7 días post-inyección no se observó banda correspondiente a ceramida, pero fue observable una banda con RF 0.65 correspondiente a cerebrósido, metabolito de la ceramida. A 30 y 150 días post inyección se observa una banda fluorescente de Rf igual a 0,80 correspondiendo a ceramida.
Genes Fas y Fas-L (Figura 3): No se encontraron mutaciones en ninguno de los tiempos estudiados en las regiones amplificadas del gen Fas- L (ligando).
Con respecto al gen fas el exón 3 no presentó mutaciones a ninguno de los tiempos estudiados pero el exón 4 si presentó las siguientes mutaciones.
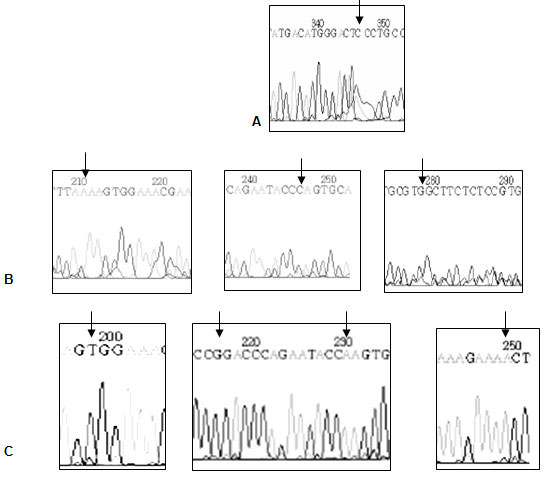
A los 7 días postinducción, se produjo un cambio de base en 347, ACTTCC cambia a ACTCCC (cambia T por C) (Figura15.3). Esta mutación no afecta al aminoácido, por lo tanto no se modificaría el producto proteico final y representa una mutación silenciosa.
A los 30 días se encontraron tres mutaciones con sentido erróneo (missense mutation, transversional), en 211 TAGAA cambia a TAAAA (cambia G por A), lo que correspondería a un cambio de un aminoácido en la proteína (ácido glutámico -> lisina); en 247 CCAAG cambia a CCCAG (cambia A por C), correspondiendo a un cambio lisina -> glutamina y en 279 GTTGC cambia a GTGGC (T por G) cambiándose asparagina -> valina. (Figura 16.3).
A los 150 días se encontraron 5 mutaciones en sentido erróneo (Figura 17.3) : en 199 TAGAA cambia a TATAA (cambia G por T) cambia el aminoácido leucina por isoleucina; en 203 AGTGG cambia a AGAGG (cambia T por A) cambia el aminoácido histidina por leucina; en 217 GCACC cambia por GCGCC (cambia A por G), cambia el aminoácido tripsina por arginina; en 231 cambia AATAC por AAAAC (cambia T por A) cambia el aminoácido leucina por fenilalanina; en 250 AAGAA cambia a AAAAA (cambia G por A) cambia el aminoácido leucina por fenilalanina.
Los productos de amplificación de los controles de ambos genes estudiados no presentaron mutaciones.
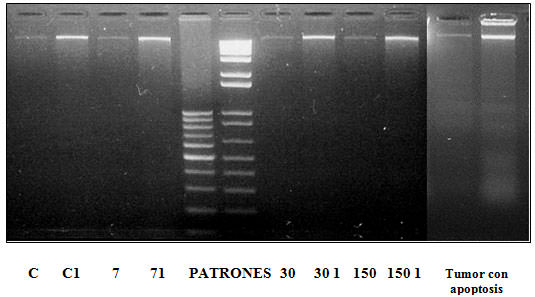
Apoptosis (Figura 4): ninguna de las muestras estudiadas mostró el patrón característico de apoptosis.
Figura 1
Glándula parótida de rata. A: Sin modificaciones ( aumento 200). B: Vacuolas en el citoplasma de las células acinares y conductos. Duplicación de capas celulares en los conductos (*) (aumento 400). C: crecimiento tumoral de tipo quístico; acúmulos de conductos intralobulillares sin presencia de acinos (*) (aumento 100). D: zona de necrosis por coagulación(*) (aumento 200). E: vacuolas en células acinares e hiperplasia epitelial ducta (*) (aumento 400) ; F :con inmunomarcación (-) (aumento 100) ; G: inmunomarcación (-) (aumento 200); H: 20% de los conductos la glándula parótida expresaron Fas (* (aumento 400); I: inmunomarcación (-) en zona de necrosis por coagulación (aumento 400) J: 70% de inmunomarcación (+)(*) (aumento 100). Figura 2
Cromatografía en capa fina, de homogenato de glándula parótida incubado con NBD-esfingomielina. Calle 1: control; calle 2: 7 días postinducción; calle 3: 30 días postinducción; calle 4: 150 días postinducción. La NBD esfingomielina aparece como una línea simple a un RF =0,15, en todos las muestras sembradas. La ceramida unida al fluoróforo aparece como una línea simple a un RF=0,80 solamente en las calles sembradas con las muestras de 30 y 150 días postinducción. Figura 3
Mutaciones del exón 4 del gen Fas. A: a los 7 días post inducción, cambio de base en 347. El cambio no produce cambio en el aminoácido codificado. B: a los 30 días post inducción, cambio de base en 211 cambia G por A, cambia ácido glutámico a lisina; en 247 cambia A por C cambia, lisina a glutamina; en 279 cambia T por G, cambia asparagina a valina C: A los 150 días postinducción, en 199 cambia G por T, cambia leucina por isoleucina; en 217 cambia A por G cambia tripsina por arginina; en 231 cambia T por A, cambia leucina fenilalanina; en 250 cambia G por A, cambia leucina por fenilalanina. Figura 4
Técnica de ADN en escalera. Fragmentación del ADN de las muestras control y problemas.C.7, 30 y 150 calles sembradas con 10?l de muestras. C1, 71, 301 y 150 1 calles sembradas con 20 ?l de muestras.En ningún caso se observa apoptosis en un porcentaje mayor al 20%. DISCUSIÓN
En este trabajo se desarrolla por primera vez un modelo experimental de inducción de la tumorogénesis en parótida de rata, utilizando el carcinógeno DMBA, un HPA (hidrocarburo policíclico aromático). Este carcinógeno ha sido ampliamente utilizado para inducir tumores en diferentes tejidos (28) entre ellos la mucosa oral de hámsteres (29, 30) y la glándula submandibular de ratas y ratones (29, 31). Los resultados de este trabajo indican que el DMBA fue igualmente efectivo para inducir provocó modificaciones en la glándula parótida.
A los 7 días postinducción del carcinógeno, se encontraron cambios histológicos leves, en un 15 % de la glándula, mientras que a nivel de ADN solo se detectó una mutación puntual que no produciría cambios en el aminoácido codificado (silent mutation). A los 30 días las modificaciones histológicas fueron severas con la presencia de zonas de necrosis por coagulación y zonas tumorogénicas, de tipo quístico .Esto se correlaciona con la aparición de 3 mutaciones en el exón 4 del gen Fas, que en todos los casos implicaron cambios en los aminoácidos codificados (missense mutation). A los 150 días se observaron importantes modificaciones histológicas, especialmente en los acinos correlacionado con la aparición de 5 mutaciones puntuales en el exón 4 del gen que al igual que las mutaciones citadas en el párrafo anterior, también modificaron al aminoácido codificado (missense mutation). Se conoce que el gen fas codifica para las proteínas de membrana Fas-receptor que al unirse a su ligando Fas-L, induce la apoptosis celular, a través de una cascada de reacciones que involucran moléculas tales como FADD, que deben unirse tanto al dominio de muerte de Fas como a una procaspasa la cual desencadena la activación en cascada de una serie de caspasas que inician la apoptosis (32). El gen fas tiene en el humano 9 exones (33), mientras que en la rata presenta 8. En tumores linfoideos malignos se encontraron mutaciones en los exones 2, 3, 4 ,6 , 7, 8 y 9 (34). En enfermedades humanas tales como el síndrome linfoproliferativo autoinmune (ALPS) (35) y en melanoma cutáneo (34), se presentan mutaciones puntuales en el exón 9 de este gen que codifica para el dominio intracitoplasmático necesario para la actividad apoptótica del receptor. Por otro lado, se ha encontrado en humanos, que en el 70 % de los casos las mutaciones que impedirían la transducción de la señal se encuentran en los exones 7 y 9 del gen fas (36) y en trabajos realizados en células tumorales de piel se demostraron una serie de mutaciones en el gen fas humano en los exones 3,5, 7 y 8, viéndose afectada la funcionalidad de la proteína en todos los casos (37). Además se observó que mutaciones de este tipo están presentes en células de linfomas, mielomas, en carcinomas renales y en cánceres de células germinales de testículo. (38,39). Se desconoce que zonas de la proteína Fas son codificadas por cada uno de los exones del gen Fas de la rata. Los resultados de nuestro trabajo, establece una relación, directa o indirecta, entre las mutaciones encontradas en el exón 4 con modificaciones en el funcionamiento de la proteína Fas, ya que en todos los casos en que se presentaron estas mutaciones se observaron modificaciones patológicas con ausencia de apoptosis.
Con el propósito de racionalizar los resultados genético-histológicos se analizó el entorno lipídico del receptor Fas especialmente la presencia/incremento del esfingolípido ceramida. El Fas-receptor es una proteína transmembrana de la familia del factor de necrosis tumoral que puede inducir la muerte celular programada a través de la unión con Fas-L. Los receptores Fas se oligomerizan en los rafts de ceramidas, los cuales a su vez se forman por el desdoblamiento de la esfingomielina de membrana por la acción de la enzima esfingomielinasa ácida, cuando la célula recibe un estímulo que induce la apoptosis (12). A nivel bioquímico, a los 7 días no se detectó la presencia de ceramida. Sin embargo se pudo observar una banda fluorescente que se correspondería con el Rf de la glucosilceramida (GlCer), un compuesto intermediario en el metabolismo de la esfingomielina. Este metabolito contiene un residuo glucosil en el C 1 de la cadena esfingoide y se encuentra aumentado en las células que no presentan apoptosis con respecto a las que si la presentan. La acumulación de GlcCer aumenta la resistencia a la apoptosis, mientras que el bloqueo de la conversión de ceramida en Glcer aumenta la sensibilidad de los tejidos a preparaciones antitumorales (40). En efecto, se ha demostrado la hidrólisis progresiva de NBD- esfingomielina a NBD- ceramida, en estudios realizados en Jurkats cells a las cuales se les indujo apoptosis a través de la vía Fas. Por el contrario en los controles tratados con esfingomielina, pero sin inducción de apoptosis se observó la aparición de, pequeñas cantidades de glucosilceramida (41). En nuestro modelo experimental, recién a los 30 y 150 días post inducción, si se pudo observarse la presencia de ceramida, disminuyendo notablemente la cantidad de la banda correspondiente a glucosilceramida. La ceramida puede inducir apoptosis por su capacidad de segregación dentro de la membrana que resulta en el arrastre de proteínas de membranas tales como el Fas-receptor. Así lo demuestran experimentos realizados incorporando ceramida exógena que estimuló la apoptosis en células tumorales. Similarmente también se observó que linfoblastos de pacientes con enfermedad de Neumann- Pick, no responden con apoptosis a la irradiación ? debido a fallas en la generación de ceramidas, pero la estimulación de fibroblastos de pacientes con esta misma enfermedad, utilizando ceramida exógena, indujo la formación de Fas-L y restituyó la sensibilidad a la apoptosis (42). Algunas células tumorales están caracterizadas por un bajo contenido de ceramidas en relación con células normales, pero en muchos tumores los niveles de ceramidas son superiores que en tejidos normales Por otra parte se ha encontrado que las ceramidas inducen no solo la apoptosis, sino también la necrosis (16). Además es conocido que se pueden desencadenar dos caminos diferentes a partir de de la inducción del receptor Fas, uno que conduce a la activación de caspasa-3 y promueve la apoptosis y otro que favorece la activación de caspasa - 8 y promueve la necrosis. (43). En nuestro trabajo a 30 días postinducción de la tumorogénesis se encontró una amplia zona de necrosis, que abarcó entre el 40 y 50% de la glándula y a 150 días las células acinares presentaron también indicios de necrosis . En ambos casos la ausencia de apoptosis determinada por DNA ladder y la presencia simultánea de ceramidas, podría indicar que la vía metabólica de la esfingomielina prosigue hacia la inducción de necrosis y no la de la apoptosis. Con respecto a la inmunocitoquímica para Fas fue negativa a los 7 días. A los 30 fue positiva en un 20 %. Se ha observado que el desarrollo inicial de algunos tumores, como los de ovario está acompañado de una alta expresión de las proteínas Fas (44), promoviendo así la apoptosis de células malignas. En este trabajo el desarrollo temprano (30 días) de los tumores también estaría acompañado de expresión alta de la proteína Fas. A 150 días la imunocitoquímica mostró un grado de inmunopositividad del 70%, correspondiendo al mismo tiempo cambios histológicos de tipo tumorogénicos y necrosis. En trabajos realizados en melanomas cutáneos malignos, se postula que la presencia de inmunoreactividad positiva para Fas, acompañada de mutaciones del gen Fas estarían involucrados en mecanismos de resistencia de las células tumorales, mediados por Fas (34).Una de las causas por las cuales las células tumorales pueden sobrevivir es porque escapan de la apoptosis inducida vía el sistema Fas- FasL, a través de mutaciones de una base en el gen Fas que modifique alguna porción de la estructura de la proteína Fas (38,39). No se conoce, en el genoma de rata los exones que codifican específicamente cada parte de la proteína Fas. No obstante la aparición de mutaciones puntuales a nivel del exón 4, las cuales probablemente no modofican su capacidad de unión a Fas-ligando, ya que la inmunomarcación con Fas-ligando fue (+) indicarían que todos los elementos de la membrana plasmática (superficie celular) necesarios para la inducción de la apoptosis estarían funcionalmente presentes. Sin embargo la ausencia de apoptosis (detectable por DNA ladder), con la presencia de modificaciones de tipo tumorigénicas sugieren que las mutaciones observadas del gen que codifica a la proteína Fas podría haber dañado segmentos relacionados con su interacción con la proteía adaptadora FADD mientras que su capacidad de interacción, a nivel de la superficie celular, con su Fas-ligando permanece funcionalmente normal. En consecuencia si bien la transmisión a través de la membrana es normal el bloqueo posterior (a nivel citoplásmico) impide la aparición del fenómeno de apoptosis. En concordancia con estos resultados se ha observado, en la formación del melanoma cutáneo maligno, que una aumentada expresión de la proteína Fas, no necesariamente predice susceptibilidad a la muerte celular programada. La apoptosis mediada por Fas puede ser bloqueada por diversos mecanismos incluida la mutación de la estructura primaria de Fas (45). También, en estudios realizados en glándula submandibular de pacientes con Síndrome de Sjögren demostraron que a pesar de una elevada expresión de Fas la apoptosis ocurrió como un evento raro, lo que podría indicar que la transmisión de la señal de muerte podría estar bloqueada (10). Por otro lado mutaciones en el DD de Fas, que formaron un DISC muy ineficiente, han sido naturalmente encontradas, tanto en ratones como en pacientes con Síndrome linfoproliferativo autoinmune (ALPS, sus siglas en inglés). (8). Las mutaciones del gen Fas en su DD conducen, por lo tanto, a la pérdida de su función como inductor de apoptosis. (37). Por otra parte la inmunocitoquímica positiva para Fas podría indicar la fragmentación funcional de Fas sugiriendo una normal asociación con el ligando pero una deficiente asociación con la proteína adaptadora FADD.
La falta de respuesta histológica, bioquímica y genómica a tiempos tempranos (7días post.inducción) sugiere que la aparición de los efectos anormales, estimulados por el carcinógeno requiere períodos más prolongados. En resumen, en este trabajo proponemos que las modificaciones genéticas del gen Fas estarían impidiendo la transducción a nivel citoplásmico, pero no a nivel de superficie celular, la señal de apoptosis, Por el contrario, probablemente las mutaciones que experimenta la proteína Fas inducen los mecanismos que estimulen la necrosis, pero no la apoptosis.
BIBLIOGRAFIA
Sanchez Torres L, Diosdado Vargas, F: Apoptosis: el fenómeno y su determinación. Téc Pecu Mex, 2003, 41(1): 49-62.
Irina Petrache I, Natarajan, V Zhen L, Medler T, Richter A, Cho Ch, Hubbard W, Berdyshev E,and Tuder R. Ceramide upregulation causes pulmonary cell apoptosis and emphysema .Nat Med. 2005 May; 11(5).
Wolf B, Green D: Suicidal tendencies apoptotic cell death by caspase family proteinases. Journal of Biological Chemistry,1999; 274: 49-52.
Schulze-Bergkamen H, Krammer P: Apoptosis in Cancer - Implications for therapy. Seminar Oncol,2004; 31 (1): 90-119.
Peter M, Legembre P, Barnhart B: Does CD95 have tumor promoting activities?. Biochimica et Biophysica Acta, 2005 ; 1755: 25-36.
Boldrini L, Loggini B, Gisfredi S, Zucconi Y, Baldinotti F, Fogli A, Simi P, Cervadoro G, Barachini P, Basolo F, Pingitore R, Fontanini G : Mutations of Fas (APO-1/CD95) and p53 genes in nonmelanoma skin cancer .J Cutan Med Surg; 2003; 7(2):112-8.
Van Den Berg A, Maggio E, Diepstra A, De Jong D, Van Krieken J, Popema M.: Germline Fas gene mutation in case of ALPS and NLP Hodgkin lymphoma. Blood, 2002; 99(4):1492-4.
Legembre P, Barnhart B,Zheng L,Vijayan S,Straus S, Puck J, Dale J, Lenardo M, Peter M: Induction of apoptosis and activation of NF-kB by CD95 require differeny signaling thresholds. EMBO reports, 2002; 5 (11):1084-1089.
Tanaka M, Suda T, Takahashi T, Nagata S: Expression of the functional soluble form of human fas ligand in activated lymphocytes, EMBO J, 1995 ; 14(6):1129-35.
Ohlsson M, Skarstein K, Bolstad Ai, Johannessen Ac, Jonsson R.: Fas-induced apoptosis is a rare event in Sjogren ' s syndrome. Lab Invest, 2001; 81(1):95-105.
Nakamura H, Koji T, Tominaga M, Kawakami A, Migita K, Kawabe Y, Nakamura T, Shirabe S, And Eguchi K: Apoptosis in labial salivary glands from Sjögren's syndrome (SS) patients: comparison with human T lymphotropic virus-I (HTLV-I)-seronegative and -seropositive SS patients.Clin Exp Immunol, 1998; 114:106-112.
Kolesnick R: The therapeutic potential of modulating the ceramide/sphingomyelin pathway. J Clin Invest, 2002; 110:3-8.
Andrieu-Abadie , lvade T. Sphingomielyn hydrolysis during apoptosis. Biochim. Biophys Acta; 2002; 1585: 126-134
Levade T,Malagarie-Casanave S, Gouaze V, Segui B, Tardy C, Betito S, Andrieu-Abadie N, Cuvillier O. Ceramide in apoptosis: a revisited role. Neurochem Res, 2002; 27(7-8): 601-7.
Reynolds A C, Maurera B, Kolesnickb R.. Ceramide synthesis and metabolism as a target for cancer therapy. Cancer Letters, 2002; 206: 169-180.
Dyatlovitskaya E, Kandyba A: Role of biologically active sphingolipids in tumor growth. Biochesmistry, 2006 ; 71(1): 17-25.
Bilderback Tr, Gazula Vr, Dobrowsky R:. Phosphoinositide 3-kinase regulates crosstalk between Trk A tyrosine kinase and p75 (NTR)-dependent sphingolipid signaling pathways. J. Neurochem, 2001; 76:1540-1551.
Zundel W, Giaccia A: Inhibition of the anti-apoptotic PI (3) K/Akt/Bad pathway by stress. Genes Dev,1998; 12:1941-6.
Reynolds A C, Maurera B, Kolesnickb R.: Ceramide synthesis and metabolism as a target for cancer therapy. Cancer Letters, 2004; 206: 169-180.
Pinti M, Troiano L, Nasi M, Monterastelli E, Moretti L, Bellodi C, Mazzacani A, Chiara M, Salvioli G, Cossarizza A: Development of real time PCR assays for the quantification of Fas and FasL mRNA levels of lymphocytes: studies on centenarians. Mechanisms of Ageing and development 2003; 124: 511-516.
Loidl A, Claus R, Deigner Hp And Hermetter A.: High-precition fluorescence assay for sphingomyelinase activity of isolated enzymes and cell lysates. J Lipids Res, 2002; 43: 815-823.
Lowry, OH, Rosembrough, A. Farr & R.J.Randall. Protein measurement with the folin phenol reagent. Journal of Biology and Chemical; 1951; 193:265-275.
Bancroft J.D.; Stevens A.; Turner D.R. Theory and Practice of Histological Techniques. New York. Churchill Livingstone. Edimburgo Melbourne. 1993
Gong, J; Traganos, F; Darzynkiewicz,Z: A selective procedure for DNA extraction from apoptotic cells applicable for gel electrophoresis and flow cytometry. Analytical Biochemistry,1994; 218,314-319.
Cryns Vl, Yuan J: Proteases to die for. Genes Dev,1998; 12:1551-1570.
Quin GP, Keough MJ. Experimental Design and Data Análisis for Biologists. Cambridge University Press. 2000
Dias M, Cabrita S, Sousa, E, Franca B, Patricio J, Oliveira C: Benign and malignant mammary tumors induced by DMBA in female Wistar rats. Eur J Gynaecol Oncol, 1999;20(4):285-8
Brunotto M, Barra J, Zarate A, Malberti A, Calderón O, Crosa M.: Oral mucosa alterations related to experimental oncogenesis of submandibular gland in rat .Acta Odontológica Latinoamericana , 2006; 19- (1). 13-21.
Cheny.K., Hsues.S, Lin L.M.: Immunohistochemical demonstration of p63 in DMBA-induced hamster buccal pouch squamous cell carcinogenesis .Oral Diseases, 2003; 9: (5) 235 -40.
Actis A.B, Lopez C, Joekes S, Eynard A.R: N-3, N-6 and N-9 dietary fatty acids modulate the growth parameters of murine salivary gland tumors induced by dimethyl benzanthracene. Prostaglandins Leukot Essent Fatty Acids, 1999; 61: 259-265.
Cheng J, Liu C, Koopman Wj And Mountz Jd.: Characterization of human Fas gene. Exon/intron organization and promoter region The Journal of Immunology; 1995,154, ( 3): 1239-1245.
Shin M, Park W, Kim S, Kang, Song K , Park J , Dong S,Pi H , Oh R, Lee J, Yoo N, And Lee S: Alterations of Fas (Apo-1/CD95) Gene in Cutaneous Malignant Melanoma Am J Pathol., 1999; 154(6): 1785-1791.
Straus S, Snellerm, Lenardom, Puck J, Stroberw: An Inherited Disorder of Lymphocyte Apoptosis: The Autoimmune Lymphoproliferative Syndrome. Ann Intern Med,2001; 130:591-601.
Tighe P, Stevens S, Dempsey S, Ledeist F, Rieux L And Edgar J. : Inactivation of Fas genes by Alu insertion: retrotransposition in an intron causin splicing variation and ALPS Genes & Inmunity,2002; 3(1); 566-570.
Lee Sh, Shin Ms, Kim Hs: Somatic mutations of Fas( APO1/DD95) gene in cutaneus squamous cell carcinoma arising from a burn scar. J. Invest. Dermatol, 2000; 114: 122-126.
Takayama H, Takakuwa T, Tsujimoto Y, Nonomura N, Okuyama A, Nagata S Andaozasa K. (): Frequent Fas gene mutations in testicular germ cell tumors. American Journal of pathology, 2002;161 (2):635-641.
Takayama H, Takakuwa T, Tsujimoto Y, Nonomura N, Okuyama A, Aozasa K: Analysis of Fas gene mutations on laser capture microdissected specimens from renal cell carcinoma. Jpn J Cancer Res. ; 93(11):1201-6. Jpn J Cancer Res, 2002. ; 93(11):1201-6.
Dyatlovitskaya E, Kandyba A: Role of biologically active sphingolipids in tumor growth. Biochesmistry, 2006, 71(1): 17-25.
Tepper A, Rurs P, Wiedmer T, Sims,P; Borst J; And Van Blitterswijk, W. (). Sphingomyelin hydrolysis to ceramide during the execution phase of apoptosis results from phospholipid scrambling and alters cell-surface morphology. The Journal of Cell Biology, 2000; 150 (1): 155-164.
Herr I, Dagmar W, Bohler, T, Angel P, Debatin K.:. Activation of CD95 (APO-1/Fas) singailing by ceramide mediates cancer therapy-induced apoptosis. The EMBO Journal,1997; 16 (20): 6200 - 6208.
Rojas P, Torres V , Leyton L And. Quest A: Caspase-dependent initiation of apoptosis and necrosis by the Fas receptor in lymphoid cells: onset ofnecrosis is associated with delayed ceramide increase Journal of Cell Science, 2002; 115, 4671-4683.
Van Haaften-Day C, Russell P, Davies S, King N, Tattersall Mh: Expression Fas and FasL in human serous ovarian epithelial tumousr. Human pathol, 2003;34(1): 74-9.
Boldrini L, Loggini B, Gisfredi S, Zucconi Y, Baldinotti F, Fogli A, Simi P, Cervadoro G, Barachini P, Basolo F, Pingitore R, Fontanini G: Mutations of Fas (APO-1/CD95) and p53 genes in nonmelanoma skin cancer .J Cutan Med Surg , 2003; 7(2):112-8.